べん毛形成制御機構の研究
はじめに
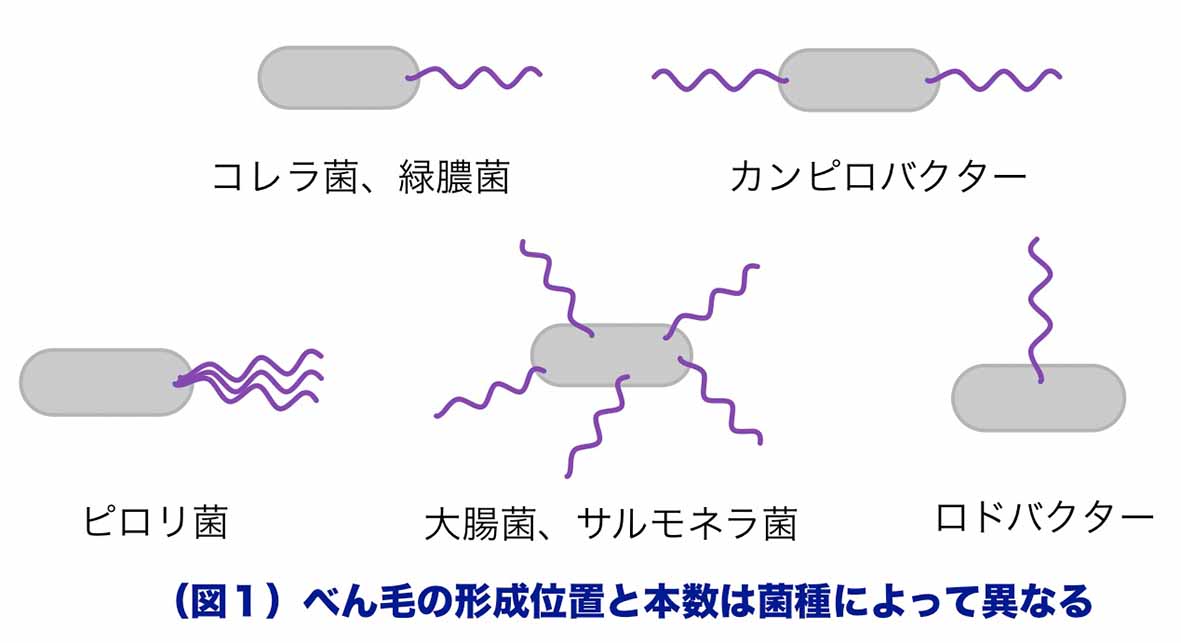 細菌の運動器官であるべん毛は、生息環境において効率よく運動できるようにその本数と位置が菌種によって厳密に制御されています(図1)。運動能低下を指標にべん毛本数が異常な変異株を選択できること、大きな構造物であるために顕微鏡でべん毛構造を確認すると同時に構成要素の局在をGFPで可視化し解析できることから、べん毛は「設置場所と数をどうやって決めるのか」という蛋白質適量配所機構を調べる上で優れた研究対象といえます。
細菌の運動器官であるべん毛は、生息環境において効率よく運動できるようにその本数と位置が菌種によって厳密に制御されています(図1)。運動能低下を指標にべん毛本数が異常な変異株を選択できること、大きな構造物であるために顕微鏡でべん毛構造を確認すると同時に構成要素の局在をGFPで可視化し解析できることから、べん毛は「設置場所と数をどうやって決めるのか」という蛋白質適量配所機構を調べる上で優れた研究対象といえます。
べん毛の数と位置を制御するFlhFとFlhG
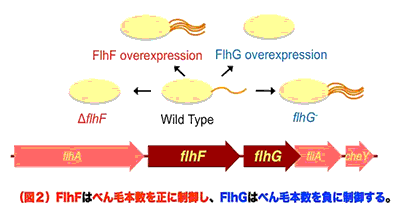 私たちは、ビブリオ菌が極に1本だけべん毛を形成する仕組みにFlhFとFlhGが関与することを明らかにしてきました(図2)。FlhFが欠損するとべん毛の配置がランダムに起こり、FlhFの過剰発現はべん毛の本数を増加させるため、FlhFはべん毛の形成位置決定だけでなく、本数の制御にも関与していることがわかっています。一方、FlhFの下流の遺伝子産物であるFlhGが欠損すると、極に複数本のべん毛が形成され、逆に過剰発現すると本数が減少します。従って、べん毛の本数はFlhFで正に、FlhGで負に制御されています。
私たちは、ビブリオ菌が極に1本だけべん毛を形成する仕組みにFlhFとFlhGが関与することを明らかにしてきました(図2)。FlhFが欠損するとべん毛の配置がランダムに起こり、FlhFの過剰発現はべん毛の本数を増加させるため、FlhFはべん毛の形成位置決定だけでなく、本数の制御にも関与していることがわかっています。一方、FlhFの下流の遺伝子産物であるFlhGが欠損すると、極に複数本のべん毛が形成され、逆に過剰発現すると本数が減少します。従って、べん毛の本数はFlhFで正に、FlhGで負に制御されています。
FlhFとFlhGの細胞内での局在
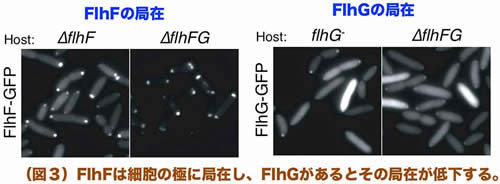
GFPを融合させたFlhFとFlhGを用いて局在を調べたところ、FlhFとFlhGは共に極に局在する性質を持ちますが、FlhFの極局在はFlhG非存在下でより顕著に観察されました(図3)。また、免疫沈降実験によってFlhFとFlhGの相互作用が検出されたことから、我々は「極局在するFlhFの分子数をFlhGが適切に抑制することで、極べん毛数を1本に制御している」とする作業仮説を立てています。
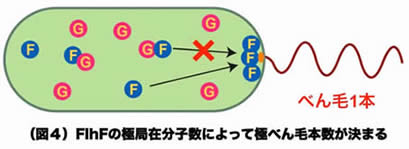
ビブリオ菌のべん毛本数・位置制御機構のモデル
FlhFは、合成直後の分泌蛋白質を細胞膜上のタンパク質輸送装置へターゲティングするFtsY/Ffhと相同性を示し、同じシグナル認識粒子(SRP)系GTPaseに分類されることから、べん毛の本数・位置の制御は、蛋白質輸送系と共通の機構で行われる可能性があります。最近他の菌種において、FlhFはGTPase活性を持つことが報告され、特にキャンピロバクターでは、GTPase活性の低下したFlhFを発現すると、べん毛本数と形成位置が異常になることから、FlhFのGTPase活性はべん毛本数・形成位置制御に重要な役割を果たしていると考えられています。一方、FlhGは細菌細胞の分裂面形成を制御するMinDとホモロジーがあります。MinDはN末端側に特徴のあるATP結合モチーフをもつATPaseであり、ATPase活性はMinDの働きに必須であることが知られています。
これまでの結果をまとめてモデルにすると、決まった数のFlhFが極に局在することでべん毛形成が1本に制御され、その過程にはFlhGの極局在が関与していると考えられます(図4)。FlhFのGTPase活性とFlhGのATPase活性は、べん毛本数・位置制御機構にどう関わるのでしょうか? また、細胞の生育段階およびべん毛の形成過程において、いつ、何分子のFlhFやFlhGが極局在するのでしょうか? 私たちは、変異体を用いた生化学・生物物理学的解析から、極べん毛本数制御の実体を明らかにしようとしています。